Transporte activo
Gradientes electroquímicos y el potencial de membrana. Transporte activo primario y secundario. Bomba Na+/K+.
Introducción
El transporte pasivo es una gran estrategia para el movimiento de moléculas dentro o fuera de una célula. Es barato, es fácil y todo lo que la célula debe hacer es quedarse allí y dejar que las moléculas se difundan a su interior. Pero... esto no funciona siempre. Por ejemplo, supongamos que el azúcar glucosa está más concentrado dentro de una célula que fuera. Si la célula necesita más azúcar para satisfacer sus necesidades metabólicas, ¿cómo puede lograr que entre ese azúcar?
Aquí, la célula no puede importar glucosa gratis mediante la difusión, porque la tendencia natural de la glucosa será difundirse hacia afuera en lugar de fluir hacia dentro. La célula debe traer más moléculas de glucosa mediante transporte activo. En el transporte activo, a diferencia del pasivo, la célula gasta energía (por ejemplo, en forma de ATP) para mover una sustancia contra su gradiente de concentración.
Aquí, veremos con más detalle los gradientes de moléculas que existen a través de las membranas celulares, cómo pueden ayudar en el transporte o complicarlo, y cómo los mecanismos de transporte activo permiten a las moléculas moverse contra sus gradientes.
Gradientes electroquímicos
Ya hemos visto los gradientes de concentración simples, en los que una sustancia se encuentra en diferentes concentraciones en una zona o en lados opuestos de una membrana. Sin embargo, debido a que los átomos y moléculas pueden formar iones y tener cargas eléctricas positivas o negativas, también puede existir un gradiente eléctrico o diferencia de cargas a través de una membrana plasmática. De hecho, las células vivas normalmente tienen lo que se llama un potencial de membrana, una diferencia en el potencial eléctrico (voltaje) a través de su membrana celular.

Crédito de la imagen: imagen de OpenStax, Biología, originalmente de Synaptitude/Wikimedia Commons
Siempre que hay una separación neta de cargas en el espacio existe una diferencia de potencial eléctrico. En el caso de una célula, las cargas positivas y negativas están separadas por la barrera de la membrana celular, con una mayor cantidad de cargas negativas al interior de la célula que al exterior de la misma. El potencial de membrana de una célula típica es de -40 a -80 milivoltios, donde el signo menos significa que el interior de la célula es más negativo que el exterior. La célula debe mantener activamente este potencial de membrana y veremos cómo se forma en la sección de la bomba sodio-potasio (a continuación).
Como ejemplo de cómo el potencial de membrana puede afectar el movimiento de los iones, echemos un vistazo a los iones sodio y potasio. En general, el interior de una célula tiene una mayor concentración de potasio (K) y una menor concentración de sodio (Na) que el líquido extracelular a su alrededor.
- Si los iones de sodio están afuera de una célula, tenderán a moverse hacia adentro de ella, de acuerdo tanto con su gradiente de concentración (la concentración de Na es más baja dentro de la célula) como el voltaje a través de la membrana (la carga más negativa está al interior de la membrana).
- Debido a que el K es positivo, el voltaje a través de la membrana provocará su movimiento hacia adentro de la célula, pero su gradiente de concentración tiende a llevarlo hacia afuera de la misma (hacia la región de menor concentración). Las concentraciones finales de potasio en los dos lados de la membrana serán un balance entre estas dos fuerzas opuestas.
A la combinación entre el gradiente de concentración y el voltaje que afecta al movimiento de un ion se le llama gradiente electroquímico.
Transporte activo: moverse en contra de un gradiente
Para transportar una sustancia en contra de un gradiente electroquímico o de concentración, la célula debe utilizar energía. Los mecanismos de transporte activo justamente hacen eso: gastan energía (a menudo en forma de ATP) para mantener las concentraciones correctas de iones y moléculas en las células vivas. De hecho, las células ocupan mucha de la energía obtenida en el metabolismo para mantener en funcionamiento los procesos de transporte activo. Por ejemplo, la mayoría de la energía de un glóbulo rojo se usa para mantener los niveles internos de sodio y potasio que difieren de los de su entorno.
Los mecanismos de transporte activo pueden dividirse en dos categorías. El transporte primario activo utiliza directamente una fuente de energía química (p. ej., ATP) para mover las moléculas a través de una membrana contra su gradiente. Por otro lado, el transporte secundario activo (cotransporte) utiliza un gradiente electroquímico, generado por el transporte activo, como fuente de energía para mover moléculas contra su gradiente y, por lo tanto, no necesita directamente una fuente de energía química, como el ATP. A continuación, veremos cada tipo de transporte activo con mayor detalle.
Transporte activo primario
Una de las bombas más importantes en las células animales es la bomba sodio-potasio, que transporta Na hacia afuera de las células y K hacia adentro de ellas. Dado que el proceso de transporte utiliza ATP como fuente de energía, se considera un ejemplo de transporte activo primario.
La bomba sodio-potasio no solo mantiene las concentraciones correctas de Na y K en las células vivas, sino que también desempeña una función importante en la generación de voltaje a través de la membrana celular en las células animales. Bombas como esta, que participan en el establecimiento y mantenimiento de los voltajes de membrana, se llaman bombas electrógenas. La bomba electrógena primaria en plantas es la que bombea iones de hidrógeno (H) en lugar de sodio y potasio.
El ciclo de la bomba de sodio-potasio
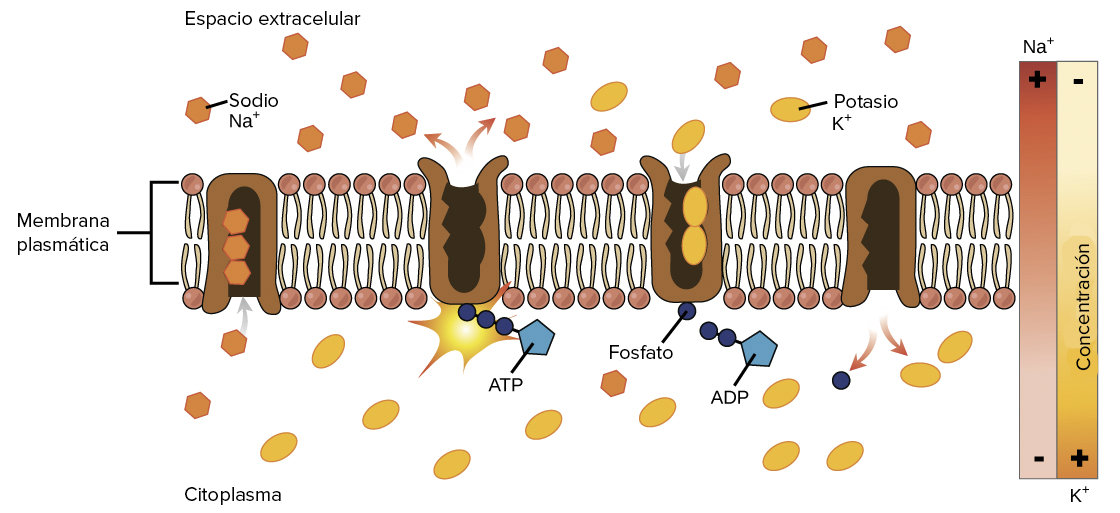
Crédito de la imagen: OpenStax Biología. Imagen modificada de una obra original de Mariana Ruiz Villarreal
La bomba sodio-potasio transporta sodio hacia afuera de la célula y potasio hacia adentro de la misma en un ciclo repetitivo de cambios de conformación (forma). En cada ciclo, tres iones de sodio salen de la célula y entran dos iones de potasio. Este proceso se lleva a cabo en los siguientes pasos:
- En su forma inicial, la bomba está abierta hacia el interior de la célula. En esta forma, realmente le gusta unirse (tiene una alta afinidad) a los iones sodio y tomará hasta tres de ellos.
- Cuando se unen los iones sodio, hacen que la bomba hidrolice (degrade) ATP. Un grupo fosfato del ATP se une a la bomba, es decir, la fosforila. En el proceso se libera ADP como producto secundario.
- La fosforilación hace que la bomba cambie de forma, reorientándose a sí misma de manera que abre hacia el espacio extracelular. En esta conformación, a la bomba ya no le gusta unirse a los iones sodio (tiene una afinidad baja por ellos), por lo que los tres iones de sodio son liberados fuera de la célula.
- En su forma orientada hacia el exterior, la bomba cambia lealtades y ahora le gusta unirse a iones de potasio (tiene alta afinidad por ellos) . Se unirá a dos iones de potasio, lo que desencadena la eliminación del grupo fosfato unido a la bomba en el paso 2.
- Sin el grupo fosfato, la bomba regresa a su forma original, y se abre hacia el interior de la célula.
- En su forma orientada hacia el interior, la bomba pierde interés en los iones potasio (tiene baja afinidad por ellos), por lo que libera los dos iones de potasio en el citoplasma. La bomba está nuevamente como en el paso 1 y el ciclo puede comenzar otra vez.
Esto puede parecer un ciclo complicado, pero solo implica que la proteína va y viene entre dos formas: una forma orientada hacia el interior con una gran afinidad por el sodio (y poca afinidad por el potasio) y una forma orientada hacia el exterior con una afinidad elevada por el potasio (y baja afinidad por el sodio). La proteína puede alternar entre ambas formas mediante la adición o eliminación de un grupo fosfato, que a su vez es controlado por la unión de los iones transportados.
Cómo genera un potencial de membrana la bomba de sodio-potasio
¿Cómo exactamente establece la bomba sodio-potasio un voltaje a través de la membrana? Es tentador responder esta duda con base en la estequiometría: por cada tres iones de sodio que se mueven hacia fuera, solamente dos iones de potasio se mueven hacia dentro, por lo que el interior de la célula es más negativo. Aunque esta proporción de cargas sí provoca que el interior de la célula sea levemente más negativo, en realidad solo representa una pequeña fracción del efecto de la bomba sodio-potasio en el potencial de membrana.
Por otro lado, la bomba sodio-potasio actúa principalmente al acumular una alta concentración de iones potasio dentro de la célula, lo que hace muy pronunciado al gradiente de concentración del potasio. El gradiente es tan pronunciado que los iones de potasio saldrán de la célula (a través de canales), a pesar de una creciente carga negativa en el interior. Este proceso continúa hasta que el voltaje a través de la membrana sea lo suficientemente alto para compensar el gradiente de concentración del potasio. En este punto de equilibrio, el interior de la membrana es negativo respecto al exterior. Este voltaje se mantendrá siempre y cuando la concentración del K en la célula se mantenga alta, pero desaparecerá si deja de importarse el K.
Para una explicación más detallada de cómo se establece el voltaje a través de la membrana, echa un vistazo al artículo potencial de membrana en la sección de neurobiología.
Transporte activo secundario
Los gradientes electroquímicos creados mediante transporte activo primario almacenan energía, que puede liberarse a medida que los iones se mueven otra vez por sus gradientes. El transporte activo secundario utiliza la energía almacenada en estos gradientes para mover otras sustancias contra sus propios gradientes.
Por ejemplo, supongamos que tenemos una alta concentración de iones de sodio en el espacio extracelular (gracias al gran esfuerzo de la bomba sodio-potasio). Si alguna ruta, como una proteína de canal o transportadora, está abierta, los iones de sodio se moverán por su gradiente de concentración y regresarán al interior de la célula.
En el transporte activo secundario, el movimiento de los iones de sodio a favor de su gradiente se acopla al transporte de otras sustancias en contra de su respectivo gradiente mediante una proteína transportadora compartida (un cotransportador). Por ejemplo, en la siguiente figura, una proteína transportadora permite que los iones de sodio se muevan en el sentido de su gradiente, pero simultáneamente lleva una molécula de glucosa en contra de su gradiente y hacia la célula. La proteína transportadora utiliza la energía del gradiente de sodio para transportar moléculas de glucosa.

Imagen modificada de "Transporte activo: Figura 4", de OpenStax College, Biología (CC BY 3.0) y "Esquema del transporte secundario", por Mariana Ruiz Villareal (dominio público)
{kind=link}
En el transporte activo secundario, las dos moléculas transportadas pueden moverse en la misma dirección (es decir, hacia la célula) o en direcciones opuestas (es decir, una hacia adentro y otra hacia fuera de la célula). Cuando se mueven en la misma dirección, la proteína que las transporta se llama simportador; si se mueven en direcciones opuestas, se llama antiportador.

Diagrama simple de un simporte (transporta dos moléculas en la misma dirección) y un antiporte (transporta dos moléculas en direcciones opuestas).
Imagen modificada de OpenStax, Biología. Imagen original de Lupask/Wikimedia Common.
No hay comentarios:
Publicar un comentario